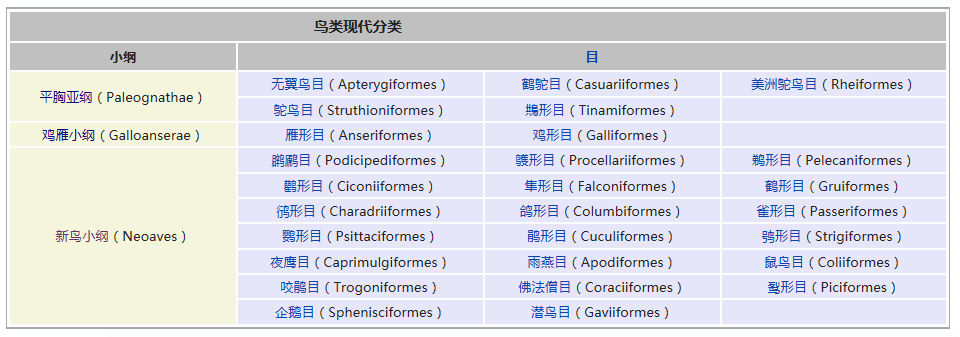
侧生动物
侧生动物域: 真核生物域 Eukaryota
界: 动物界 Animalia
亚界: 侧生动物亚界 Parazoa
Grant & Todd, 1838
门
多孔动物门 Porifera
侧生动物是动物界里的一个古老的亚界,和其领鞭毛虫的祖先不同,它们有宏观的结构及分化的细胞,但又不像真后生动物一样有组织。现在唯一留活着的侧生动物只有海绵,属于多孔动物门。有些也会包括进扁盘动物门,此门只有丝盘虫同一个物种。侧生动物是不对称的,而其他所有的动物则总是会有一些的对称。侧生动物现在包含有五千多个物种,约有150种生活在淡水里。
多孔动物门界: 动物界 Animalia
亚界: 侧生动物亚界 Parazoa
门: 多孔动物门 Porifera
Grant in Todd, 1836
纲
钙质海绵纲 Calcarea
六放海绵纲 Hexactinellida
寻常海绵纲 Demospongiae
层孔虫纲 Stromatoporata
多孔动物门(学名:Porifera)为原始的多细胞生物,也称海绵动物门(Spongiatia或Spongia),一般称为海绵。海绵没有神经、消化和循环系统,相反,它们大多依靠海水流过自己的身体来获得食物和氧气并消除废物,就像海底中的滤水器。
由于形状像植物,而且根植在海底中,所以海绵千百年来都被认为是植物,但后来发现它们几乎具备所有最基本的动物特征,所以到十九世纪中期才被归入动物界里头。但近期基于基因组资讯与系统发生学的更多证据,所以应否把它们保留在动物界还是独立出来自成一个界别的论点在生物学界仍在不断讨论中。
海绵动物体壁上有很多小孔(入水孔),游离的一端有大孔开口。
海绵的体型多种多样,小的不过几克,大的却有45公斤,最大的海绵跨度能有4.2米。海绵的颜色同样是丰富多彩。
细胞虽已开始分化,但未形成组织和器官,也没有形成真正的胚层(见内胚层、中胚层或外胚层);
体壁由内、外两层细胞构成。外层细胞扁平,称皮层(扁细胞层);内层细胞称胃层(襟细胞层),生有鞭毛,多数具有原生质领,称“领细胞”(choanocytes),主要行摄食和细胞内消化的作用;入水孔通入体内的沟道,与领细胞组成鞭毛室和出水口组成复杂的沟道系统。含有食饵的海水,由于内层细胞鞭毛的不断振动,从入水孔流入体内,不消化的东西随着海水从顶端的出水口排出体外。
在内、外两层细胞之间,还有一层中胶层,其中有像变形虫的游离细胞、生殖细胞、造骨细胞、海绵丝细胞等。
海绵动物体壁内多具有支持的针状骨骼,称内骨骼(Endoskeleton),由胶原蛋白(collagen)、碳酸钙(CaCO3)和二氧化硅(SiO2)组成。
海绵没有神经系统,但海绵细胞共同捕食、分工消化,所以被认为是动物界器官形成的开始。
大多为雌雄同体(Hermaphroditic)
分类
钙质海绵纲 (Calcispongea)
六放海绵纲 (Hexactinellida)
寻常海绵纲 (Demospongiae)
层孔虫纲 (Stromatoporata)
旧分类中,多孔动物门可分为下面一些纲:
普通海绵纲 (Demospongea)
玻璃海绵纲 (Hyalospongea)
钙质海绵纲 (Calcispongea)
硬海绵纲 (Sclerospongea)
中生动物门中生动物是一种多肉海洋无脊椎寄生动物,现今依然不清楚它们是退化了的扁形动物,还是独立发展出的。一般而言,这些细小、难以理解的生物是由包附在一个或多个生殖细胞上的纤毛种皮细胞所组成的。数十年前,中生动物被分类为一个门,但分子种系发生学的研究却显示这类谜样的中生动物其实是多系群的,亦即,它们是由两个不相关的类群所组成的。
中生动物有着复杂的生命周期,会在无性和有性的阶段间交替。在头足类动物的肾内有许多类似虫一般,由固定数量的体细胞组成的生物(线形体)。在线形体的核心内有轴细胞,这些细胞会产生新的线形体。之后,线形体再转变成菱形体,以准备性发展。性发展起始于菱形体的轴细胞中一个称做 infusorigen 的区域内,在此处发展出阿米巴状的精子来。套细胞会转变成卵。精子由轴细胞中出去,并和卵结合而形成滴幼虫,然后幼虫会爬出来。最后,章鱼再次被年轻虫状的动物感染。
中生动物一度被认为是原生生物和动物之间演化的一个过渡型态,但现在,它们则被认为是退化了或简化了的动物。它们的纤毛幼虫和吸虫的纤毛幼虫相似,且它们的内部增殖也和在吸虫体内发生的现象相似。中生动物 的 DNA 有较低的GC含量(40%)。此一数值和纤毛虫相似,但纤毛虫通常有双细胞核。其他和中生动物相关的还包括环节动物、纽形动物和真涡虫等类群。
中生动物有两个主要的类群:菱形动物门和直泳动物门,有时也包括扁盘动物门和单胚动物门。
单胚动物门只包含一个只在十九世纪时有过一次描述的物种,且自此便不曾再见过。因此,许多人怀疑它们是一个真实的类群。根据描述,这种动物只有单一层的组织。
菱形动物门
菱形动物门发现于头足纲的原肾管内。它们以二十至三十个细胞排成数毫米的长度,这些细胞包括前端的边缘巢房和一个长条状称之为轴细胞的生殖细胞。轴细胞可以无性地发展成虫状的幼体,也可以生成卵和精子,以自体受精来产生纤毛滴幼虫。
菱形动物门有三个属:二胚虫属、 Pseudicyema 和 Dicyemennea 。
直泳动物门
直泳动物门发现于许多海洋无脊椎动物的身体间隙内,如组织间隙、生殖腺、气囊等。此类病原体会导致不同物种的宿主丧失生殖能力。
最知名的直泳虫门动物为蛇尾的寄生虫。
真后生动物界: 动物界 Animalia
亚界: 真后生动物亚界 Eumetazoa
Butschli, 1910
分支
辐射对称动物 Radiata
两侧对称动物 Bilateria
真后生动物(学名:Eumetazoa)是指所有具有细胞组织的动物。动物中只有侧生动物和中生动物中的三个门不属于真后生动物。
真后生动物细胞间有连接,就是所谓的"紧密连接"。它们的胚胎最少会发生两个胚层:内胚层和外胚层。只有真后生动物才具有特化的细胞类型如神经细胞。
真后生动物可再分为两侧对称动物和辐射对称动物。后者包括刺胞动物 (Cnidaria)和栉水母动物门 (Ctenophora)。但这种分类并不能反映真实的种系发生。按照亲缘分支分类法,栉水母动物和两侧对称动物有更多的亲缘关系。
两侧对称动物界: 动物界 Animalia
亚界: 真后生动物亚界 Eumetazoa
演化支: 两侧对称动物 Bilateria
演化支
后口动物 Deuterostomia
原口动物 Protostomia
两侧对称动物(学名Bilateria),又名左右对称动物,与辐射对称动物(Radiata)共同组成真后生动物(Eumetazoa)。
两侧对称动物的身体由三层胚层发育而来,即拥有中胚层。三胚层是由两胚层演化而来的。故两侧对称动物比辐射对称动物高级。
顾名思义在体形上它们呈两侧对称。在躯干前段开始形成脑部(Cephalisation)。某些软体种类环状肌肉(收缩会导致该处躯干收窄)在外,纵行肌(收缩导致该处躯干复原)在内。它们大部分具有长的肠脏和血液系统。在表皮(Epidermis)和肠之间有体腔(Coelom),它对内部会填充体液,形成静水压保持其形态。若果存在血液系统,则动物的排泄会通过原肾和后肾完成。
它们有三种卵裂方式:螺旋(原口动物Protostomia),辐射(后口动物Deuterostomia)和两侧对称(原口动物,后口动物)。
后口动物界: 动物界 Animalia
亚界: 真后生动物亚界 Eumetazoa
演化支: 两侧对称动物 Bilateria
总门: 后口动物总门 Deuterostomia
Grobben, 1908
门
脊索动物门 Chordata
半索动物门 Hemichordata
棘皮动物门 Echinodermata
异涡动物门 Xenoturbellida
无腔动物门 Acoelomorpha
古虫动物门 Vetulicolia
后口动物总门(拉丁语学名:Deuterostomia)是动物中的一个总门,在真后生动物亚界的两侧对称动物分支中,和原口动物相对。后口动物和原口动物可以由胚胎发育相区分;在后口动物中,第一个开口(胚孔)变成了肛门,而在原口动物中则变成了嘴巴。
后口动物共有五个现存的门:
脊索动物门 Chordata (脊椎动物和其同类)
棘皮动物门 Echinodermata (海星、海胆和海参等)
半索动物门 Hemichordata (肠鳃,笔石可能也是)
异涡动物门 Xenoturbellida (两个外表似虫的物种)
无腔动物门 Acoelomorpha
毛颚动物门可能也属于后口动物,已灭绝的类群还包括古虫动物门。
后口动物和前口动物,其受精卵一开始都会发育成一个由细胞组成的小球,称之为囊胚。在后口动物中,初期的部分会平行或垂直极轴,这称为幅射卵割,也会出现在某些原口动物上,如触手冠动物。大多数的后口动物会行不定卵割,其细胞在胚胎中的发展不是由母细胞来决定的。因此,若将一开始的四个细胞分开,每一个细胞都能够形成一个完整的幼体,且若一个细胞被移出了囊胚,其他的细胞就会补充进来。
半索动物和脊索动物都有腮裂,且原始化石中的棘皮动物也显示出有腮裂的记号。在所有脊索动物,包括尾索动物(在幼体的阶段)之中,都有着中空的神经索。一些的半索动物也会有管状的神经索,在胚胎早期时看起来像是脊索动物的中空神经索。因为棘皮动物那退化的神经系统,分辨其祖先是否也是如此是不可能的,但依其他不同的事实来看,有相当的可能性,所有现存的后口动物都演化自一个有腮裂、中空神经索和分段的身体。此一祖先应类似寒武纪时被称做古虫动物的小后口动物类群。
原口动物界: 动物界 Animalia
亚界: 真后生动物亚界 Eumetazoa
演化支: 两侧对称动物 Bilateria
(未分级) 原口动物 Protostomia
总门
蜕皮动物总门 Ecdysozoa
冠轮动物总门 Lophotrochozoa
原口动物(拉丁语学名:Protostomia)有完整的消化道。胚胎时期的原口会发展为成体的口,而肛门则是另外形成的。蜕皮动物(Ecdysozoa)和冠轮动物(Lophotrochozoa)都属于原口动物。其卵裂方式为螺旋式,因其不均匀分裂会导致中间有一个时期,出现四个大细胞(Macromere)和四个小细胞(Micromere)。该类动物若具有体腔,则体腔是由中胚层形成的。
具体与后口动物的区别,请参看后口动物条目。
原口动物包括:
蜕皮动物(Ecdysozoa)
动吻动物门(Kinorhyncha)
鳃曳动物门(Priapulida)
线虫动物门(Nematoda)
线形动物门(Nematomorpha)
有爪动物门(Onychophora)
铠甲动物门(Loricifera)
缓步动物门(Tardigrada)
节肢动物门(Arthropoda)
冠轮动物(Lophotrochozoa)
软体动物门(Mollusca)
环节动物门(Annelida)
苔藓动物门(Bryozoa)
腕足动物门(Brachiopoda)
扁虫动物(Platyzoa)包括以下七门(或直接归类到冠轮动物)
扁形动物门(Platyhelminthes)
腹毛动物门(Gastrotricha)
颚胃动物门(Gnathostomulida)
微颚动物门(Micrognathozoa)
轮形动物门(Rotatoria)
环口动物门(Cycliophora)
棘头动物门(Acanthocephala)
蜕皮动物界: 动物界 Animalia
亚界: 真后生动物亚界 Eumetazoa
(未分级) 两侧对称动物 Bilateria
原口动物 Protostomia
超门: 蜕皮动物总门 Ecdysozoa
Aguinaldo et al., 1997
门
泛节肢动物(Panarthropoda)
节肢动物门(Arthropoda)
有爪动物门(Onychophora)
缓步动物门(Tardigrada)
Scalidophora
动吻动物门(Kinorhyncha)
鳃曳动物门(Priapulida)
铠甲动物门(Loricifera)
线虫动物门(Nematoda)
线形动物门(Nematomorpha)
蜕皮动物(Ecdysozoa)是一大类原口动物,包括节肢动物门、线虫动物门和几个小门。最初由Auinaldo等人于1997年定义,主要根据是18S 核糖体RNA树。而这个分类同时也被一系列形态学证据所支持,因为这个分类包括所有蜕掉外骨骼的动物(见蜕皮(ecdysis))。与蜕皮动物接近的定义也曾经被Perrier在1897年和Seurat在1920年仅依据形态作出。
蜕皮动物最显著的共同特征是一个三层的表皮(英文cuticle),由有机物组成,能够随着动物生长周期性蜕掉。蜕皮动物由此得名。蜕皮动物缺乏运动纤毛,产生变形虫样精子,其胚胎不像其他原口动物一样螺旋卵裂。蜕皮动物的一些分支还有其它一些共同特征,比如缓步动物和线虫动物都有一个三辐射对称的咽(pharynx)。
蜕皮动物包括如下门:节肢动物门、有爪动物门、缓步动物门、动吻动物门、鳃曳动物门、铠甲动物门、线虫动物门和线形动物门。其它一些门类,比如腹毛动物门,曾被认为可能是其中的一员,但因其缺乏蜕皮动物的主要特征,现在通常被划出。泛节肢动物的共同特征是具有分节的身体,原来以为由环节动物演变而来,而与其一起组成分节动物(Articulata)。然而它们并不具有太多的共同特征,现在看来他们分别演化出了分节的特点。此外,从18S 核糖体RNA树来看,后口动物中的毛颚动物门可能与其余后口动物不同而属于蜕皮动物。蜕皮动物中不属泛节肢动物的成员曾经被组成环神经动物(Cycloneuralia),但通常被认为是并系群。
冠轮动物界: 动物界 Animalia
亚界: 真后生动物亚界 Eumetazoa
(未分级) 两侧对称动物 Bilateria
原口动物 Protostomia
超门: 冠轮动物总门 Lophotrochozoa
冠轮动物(学名Lophotrochozoa)是动物界中的一大支,属于两侧对称动物,与蜕皮动物(Ecdysozoa)组成原口动物。原口动物和后口动物(Deuterostomia)并列为两侧对称动物的两个分支。本词由触手冠动物(Lophophorata)和担轮动物(Trochozoa)两词缩合而成。
冠轮动物包括传统上的:
软体动物门(Mollusca)
环节动物门(Annelida)
纽形动物门(Nemertea)
星虫动物门(Sipunculida)
螠虫动物门(Echiura)
须腕动物门(Pogonophora)
触手冠动物(包括苔藓动物门(Bryozoa)、内肛动物门(Entoprocta)、腕足动物门(Brachiopoda)和帚虫动物门(Phoronida))。
此外根据18S rRNA序列,轮形动物门(Rotifera)(或许仅部分轮形动物)和新近发现的环口动物门(Cycliophora)也很可能属于冠轮动物。
所有冠轮动物属于原口动物(Protostomia),即在胚胎期,口的发育在肛门发育之前形成。冠轮动物尽管形状上相差很多,但通常为蠕虫状,身体柔软。
担轮动物
传统上最大的两类担轮动物是软体动物和环节动物。担轮动物的生活史具有一些关键的共性。幼虫孵化后成为微小的游泳体,称为担轮幼虫。担轮幼虫在身体中部拥有两条纤毛带,用于游泳和捕获食物,而在“顶端”有一丛长的鞭毛。担轮动物的幼虫非常相似,尽管其成体区别很大。直到最近,人们一直认为节肢动物和环节动物是近亲,其原因是这两类动物都有身体分节的现象,但节肢动物不具有担轮幼虫,分子生物学也没有支持这一观点的证据。
触手冠动物
触手冠动物曾经被认为是单系群(Monophyly),包括帚虫动物、外肛动物和腕足动物。其中后两类拥有大量化石记录。它们都具有触手冠(Lophophore),上有中空的触手,用于捕食。曾经有人认为触手冠动物和后口动物有密切关联,但较新的研究将其与担轮动物联繋在一起。
新近的研究发现,不论担轮动物或触手冠动物都不是单系群。其中最大的软体动物和环节动物两个门的各纲在演化树上分属不同分支,尤其环节动物的多毛纲(Polychaeta)的各个分支处于冠轮动物的基部,可能为并系群(英文paraphyly),代表了冠轮动物的原始形态。而各小门可能为特化的类群,其独立性需要进一步讨论。关于冠轮动物内部各门类的演化分类有多种形态学方法以及分子生物学方法,结论尚不能确定。
扁虫动物界: 动物界 Animalia
超门: 扁虫动物总门 Platyzoa
扁虫动物原本是扁形动物门的同义词,此处是几个门的统称,属于两侧对称动物。扁虫动物总门(Platyzoa)包括以下门类,但此分类尚未成为共识:
扁形动物门 (Platyhelminthes)
腹毛动物门 (Gastrotricha)
颚胃动物门 (Gnathostomulida)
微颚动物门 (Micrognathozoa)
轮形动物门 (Rotatoria)
棘头动物门 (Acanthocephala)
环口动物门 (Cycliophora)(可能)
扁形动物和腹毛动物没有体腔。其他的门则具有假体腔。它们的颚和咽构造相似。但是行寄生生活的棘头动物没有这些结构。它们形成一个单系群——Gnathifera。
扁虫动物总门的动物和担轮动物是近亲,有时被归到后者。两者组成螺旋卵裂动物(Spiralia)。软体的扁形动物门通常为极扁平的蠕虫,包括自由生活的和寄生的种类。栖于海水、淡水和陆地多种生境,广布于世界各地。体长从1毫米的若干分之一至15米。有3个类型:涡虫、吸虫和绦虫。扁虫身体两侧对称,不分节,背腹扁平;无呼吸系统、骨骼、循环系统和体腔。器官之间的空隙为固体的结缔组织填充。
辐射对称动物界: 动物界 Animalia
亚界: 真后生动物 Eumetazoa
门
刺胞动物门 Cnidaria
栉水母动物门 Ctenophora
辐射对称动物(学名Radiata)与两侧对称动物(Bilateria)共同组成真后生动物(Eumetazoa)。
这些动物的外形为球形或桶形。
辐射对称动物身体由两层胚层构成。中间有一中胶层(Mesogloea)。
刺胞动物门界: 动物界 Animalia
(未分级) 辐射对称动物 Radiata
门: 刺丝胞动物门 Cnidaria
Hatschek, 1888
纲
珊瑚纲 Anthozoa
钵水母纲 Scyphozoa
十字水母纲 Staurozoa
立方水母纲 Cubozoa
胞内寄生水螅虫纲 Polypodiozoa
水螅纲 Hydrozoa
侧水母纲 Dipleurozoa
原水母纲 Protomedusae
水螅锥石纲(门) Hydroconozoa
刺胞动物门(Cnidaria ),又名刺丝胞动物门,是动物界的一个门。除极少数种类为淡水生活外,绝大多数种均为海洋生活,大多数在浅海,有些在深海,现存种类大约有11000种。刺胞动物曾经和栉水母动物一起分作腔肠动物门,后栉水母动物独立成一门。
刺丝胞动物门动物有如下特点:
大部分为肉食性,少部分种类获得能量来自于体内共生生物行光合作用得来的。
其躯干呈辐射对称,水生,大多固着生活
体壁有两层细胞组成,表皮和肠表皮,两者之间有一层凝胶状的中胶层,起支持作用。
有肠腔,有一围口部(Peristom),既是口也是肛门。
有超过20种的刺胞,刺丝胞中含有刺丝囊。刺丝囊一端的鬃样突起的刺针,受刺激时,激起刺丝囊排空。刺丝胞的表面有突出,胞体内有棍状结构。刺胞内有高尔机体分泌物质,在压力作用下会释放。
在肠腔中进行胞外消化
弥散的神经系统,呈网状
水母体有感觉器官,能感受光和重力
有雌雄同体或异体
水母、珊瑚、海葵、水螅都被归类为刺丝胞动物。
栉水母界: 动物界 Animalia
门: 栉水母动物门 Ctenophora
Eschscholtz,1829
纲
触手纲 Tentaculata
无触手纲 Nuda
硬骨纲 Scleroctenophora
栉水母(学名:Ctenophora),又名海胡桃,是一类两胚层动物,属辐射对称动物,现被划分为栉水母动物门。原和刺胞动物一起分在腔肠动物门,作为无刺胞亚门,现分出。
有较发达的中胶层,拥有简单的网状神经系统和肌肉系统。
两胚层;伞缘下延,向内收缩,致身体成球形、卵圆形、扁平形等,体外具有栉板排列成纵行的纤毛带8条,触手没有刺细胞;身体左右对称或呈辐射对称,胶质厚而透明,游泳时振动栉板;反口极有一平衡感觉器;没有胃丝。
生物学上,栉水母与水母为不同门,因栉水母并无刺胞。通过全基因组测序,发现栉水母是最古老的多细胞动物,而很多结构远比栉水母简单的生物(如海绵生物门,也叫多孔动物门,以及扁盘生物,这些生物都有一定的细胞分化,没有组织分化,没有形成胚层,没有神经系统,没有肌肉系统),反而与高等生物关系更近。从基因组和转录组分析,最早的多细胞生物祖先没有神经系统和肌肉系统,栉水母和水母发生了趋同进化,两者独立的进化出了神经系统。 栉水母的神经发育及运作的基础机制不同于水母。例如,水母及其他高等动物中神经系统发育关键基因在栉水母中都不存在。同样,栉水母也不使用水母及其他动物常用的神经递质,如多巴胺,乙酰胆碱,血清素等。于此同时,栉水母依赖使用谷氨酸作为最主要的神经递质,尽管其他动物同样使用谷氨酸作为神经递质,但栉水母中谷氨酸受体基因远比其他动物的多。由于其他动物的神经系统都是由原始水母的神经系统进化出来的,拥有与水母相同的基础机制,因此可以认为栉水母的神经系统是独立于其他动物的全新的神经系统。
栉水母门可以分为3个纲5个目:
触手纲(Tentaculata):有触手的水母。
球栉水母目(Cydippida):如侧腕栉水母(Pleurobrachia)。
兜水母目(Lobata):例如Mnemiopsis。
带栉水母目(Cestida):例如带栉水母(Cestum)。
扁栉水母目(Platyctenea):例如扁栉水母(Ctenoplana)。
无触手纲(Nuda):无触手的水母。
瓜水母目(Beroda):例如瓜水母(Beroe)。
硬骨纲(Scleroctenophora)
软体动物域: 真核域 Eukarya
界: 动物界 Animalia
亚界: 真后生动物亚界 Eumetazoa
(未分级) 两侧对称动物 Bilateria
原口动物 Protostomia
超门: 冠轮动物总门 Lophotrochozoa
门: 软体动物门 Mollusca
多板纲 Polyplacophora
单板纲 Monoplacophora
双壳纲 Bivalvia
掘足纲 Scaphopoda
腹足纲 Gastropoda
头足纲 Cephalopoda
喙壳纲 Rostroconchia
太阳女神螺纲 Helcionelloida
竹节石纲 Tentaculita
软舌螺纲 Hyolitha
软体动物门(学名:Mollusca)属于无脊椎动物,就其物种多样性而言,是动物界的第二大门,仅次于节肢动物门,共有十万多种。软体动物能适应许多不同环境,分布广泛,从寒带、温带到热带,从海洋到河川、湖泊,从平原到高山,陆地、淡水和咸水中都有大量成员,例如蜗牛、河蚌、海螺、乌贼等物种。
软体动物型态、习性差异甚大,最大的软体动物大王乌贼的腕展开可达12米,最小的螺类却仅有1厘米长。但是它们有共同的基本特征,身体无内骨骼且软,大多数不分节,身体结构可分为头、足、内脏团和外套膜4个部分。部分软体动物的外套膜会分泌出钙质的硬壳保护身体。外套模的形状因种类而不同。除了成年期的腹足动物之外,软体动物的的壳体都是左右对称的。
软体动物大多有壳,如田螺、文蛤等贝类;少数在陆地上的则有蜗牛、蛞蝓;章鱼、乌贼、海蛞蝓的外壳已消失;软体动物多数靠一条肉脚向前滑动,以此移动自己的身体,很多都有一个盘绕的外壳来保护蜗在里面的柔软的身体。
软体动物最主要的特征就是身体柔软,并无内骨骼,大部分呈左右对称、不分节,它们外层皮肤会自背部折皱形成外套膜包围全身,并能够分泌保护用的石灰质介壳,然而部分软体动物的外壳已退化(如蛞蝓、章鱼)或是藏至体内(如乌贼)。斧足类、腹足类的壳表还有生长线,可以看出年龄大小。软体动物的腹部有肌肉足或腕,但也有的肉足已经退化,是运动器官。
大多数软体动物的头上长有和眼睛一样的触角,用来感知周围的情况,它们有几千颗微小的牙齿,称为齿舌,是他们多数特有的器官,由多列角质齿板组成,形状类似锉刀,主要用于摄食。有栉鳃,表面具纤毛,可以激动水流,双壳类可用此过滤水中的食物颗粒。
水生的软体动物排泄器官为肾,会排出氨或尿素,它们呼吸器官为鳃,由外套膜形成位于外套与身体之间,可以从流入外套腔的水中吸取氧气。而像蜗牛等陆生的软体动物排泄器官也是肾,但是用肺呼吸,并排泄尿酸。
有些软体动物是雌雄异体;有些是雌雄同体。头足纲及部分腹足纲体外受精,雌雄同体者则异体受精。不同的软体动物有不同的产卵方式。像斧足类、石虌等把卵直接产在海水中,并无保护措施,因此产软数量极多,但能顺利发育的却不多。有些海螺则会把卵埋起来,或分泌胶质包住。而腹足类的卵大都有胶质包住或具有壳。
部分的软体动物具有经济价值,斧足类大都能产真珠。还有许多种淡水产的蚌类,分泌的真珠质可以制钮扣。鲍鱼、蛤蜊、牡蛎等可供人类食用,更有许多软体动物提供了鱼类大量食物来源。软体动物除了对人类有益,部分物种对人类有害,例如:斧足类的凿船虫,这种贝类生活在海里,会破坏船只或栖身在木洞里。还有部分淡水的软体动物,是肝蛭、日本住吸虫的中间寄主,会传染疾病。
软体动物的身体主要可分为头、足、内脏团3个部分,部分生物还具有从外套膜形成的壳。
头
头位于身体的前端。
肉足
足可能位于头后或身体腹面,是由体壁伸出的由多肌肉质组成的运动器官。
内脏团
内脏团位于身体背面,由柔软的体壁包围着内脏器官。
外套膜与外套腔
外套膜是软体动物背侧的体壁向下褶与伸展形成的,经常包裹整个内脏团,内脏囊与外套膜之间的空腔即为外套腔。外套腔是许多器官的开口,如肛门、鳃孔、肾孔、生殖孔等。双壳纲生物整个柔软生体都在扩大的外套腔内。
壳
有些软体动物的外套膜会分泌碳酸钙、几丁质和贝壳硬蛋白而形成外壳,外壳的最外层是角质层,中层则是柱状方解石,最内层则是层状方解石。大部分的软体动物壳主要由文石组成,但是它们生产的蛋壳却由方解石组成。
神经系统
除了双壳类有三条外,大部分软体动物有两对主要的腹神经索,一个主要连结到内脏团,另一个连结到肉足。两对神经索与主要的神经节都在肠道下侧。
部分软体动物有大脑,并且环绕食道。大多数软体动物具有眼睛有眼头,且有一对具传感功能的触角,可检测化学物质、感受外界的震动等。
消化系统
呼吸系统
大多数软体动物只有一对鳃,有的甚至只有一个。大部分的腮形成羽毛状,有些则成丝状。软体动物的腮可使水从底部进入顶部排出。腮的单丝有三种纤毛其单丝有三种纤毛,其中一个穿过外套膜以驱动水流,而另外两条丝可保持腮的清洁。在腮里有个东西叫嗅检器(osphradia),当它检测到有害物质或是泥沙进入外套膜时,腮的纤毛变化停止跳动,直到有害物质已消失。每个鳃都具有连结到体将的血管,与运送血液到心脏的血管。
循环系统
大部分的软体动物是开放式循环系统。虽然软体动物是三胚层动物,但是它们的体腔却相当小并包含了心脏与生殖腺。体腔内大多是血腔,能让血液与体液循环至各个器官。软件动的血液含有血蓝蛋白,与氧气结合时会产生Cu2+,因此它们的血液呈现蓝色。它们的心脏有些只有一个心房,有些有多个心房,它们从腮得到含氧血,并送到心室,然后再送到相当短的主动脉并流至全身。
软体动物的心房还有部分的排泄功能。心房过滤血液中的废物,并排到体腔中形成尿液。位于背部的一对肾管,可从尿液中提取可用的物质、排出无用的废物,经由管道排到体外。
泌尿系统
生殖系统
分类
生物学家根据软体动物的硬壳和软体结构的差异,将软体动物分成了10个纲,分别是单板纲、多板纲、无板纲、腹足纲、掘足纲、双壳纲、喙壳纲、头足纲、竹节石纲和软舌螺纲。
单板纲
在寒武纪早期时已出现,新笠贝是其代表,它们的特征是壳的形状像帽子或勺子。
多板纲
多板纲生物在寒武纪晚期已出现,它们的身体对称,是椭圆形,有8块骨板组成的背壳,因此称多板纲。
无板纲
无板纲生物并没有骨板或贝壳,因此称为无板纲生物,它们的外表带有钙质针状的角质外皮,具有保护作用。它们的身体两侧对称,像蠕虫似的。目前没有找到无板纲生物的化石,现代生物海兔是其代表。
腹足类
腹足类是软体动物中最大的一类,大约有七万五千多种,它们的肉足都是长在身体的腹面,因此被称为腹足类。除了蛞蝓、海蜗牛外,它们都有螺壳,因此又被称为螺类。
这类生物的螺壳,形状、大小、颜色会因种类不同而相异。但是螺壳上的螺纹大多数是顺时针方向,也可以从它们圆锥形的螺壳上,判断出年龄大小;凡是顶端的地方螺纹越细越紧密的,年纪就越大。
陆地常见的腹足类包含蜗牛、蛞蝓、螺等;海洋中的腹足类较多,如龙宫贝、夜光贝、鲍鱼等都是。为了适应各种环境,它们的肉足可以在许多地方爬行,并分泌出黏液来。
腹足纲从寒武纪早期就出现了,一直繁衍到现代,陆地、海洋、淡水里都有它们的分布。
斧足类
斧足类因都具有双壳因此又称双壳纲,河蚌、海扇、蛤蜊以及大部分的贝壳都属于双壳纲。它们最主要的特点就是有两个壳瓣,两个壳办之间有铰合构造;它们缺乏明显的头部,但是在腹侧有斧状的肉足因此称为斧足类。它们在寒武纪时就出现在地球上,海洋、淡水都能发现它们的存在。
头足类
头足纲最主要的特征就是具有神经高度集中的头部,是软体动物中最高级的生物,它们都是海洋中的肉食性动物。头足纲动物两侧对称,头部长在身体前方,头部两侧有发达的眼,口中有角质的颚片;口的周围环列着一圈能够用来捕食其它动物的触手或腕。他们有的具有外壳、有的隐藏至身体内部、有的甚至完全退化。地质史上,著名的头足纲生物包含鹦鹉螺类、杆石、菊石、箭石等,现在著名的生物包含章鱼、乌贼等。头足纲在寒武纪时已经出现了。
掘足类
掘足纲与斧足类有亲缘关系,它们的介壳是烟嘴状的,在发生之初仍然是二枚贝的形态,代表生物有齿贝。
它们身体呈两侧对称,且是圆柱状,具有个管状介壳。它们大多生活在海边的泥沙中,但也有生活在15,005英尺的深海中。它们的足部在前端,后多则是外套腔开口。头部不发达,口位于膨大咽头的前端,并在足部背面。消化管呈现U字状,肛门开口于外套腔。有肾管一对,开口处在肛门略后。并没有鳃,依靠外套膜呼吸。口周围有触角,有感觉作用。其神经系统发育正常,有脑、脏、足、侧等成对之神经结。它们都是雌雄异体,只有一个生殖腺,生殖细胞由右侧肾管排出之。
喙壳纲
喙壳纲是一个已经灭绝的生物纲,存在于寒武纪到奥陶纪,它们的壳看起来有两瓣,但无法铰合,形成“假双壳”形态的单壳,代表生物有海拉尔特壳等。
竹节石纲
竹节石纲是一个已经灭绝的生物纲,存在奥陶纪到泥盆纪,身体呈辐射对称,壳是单锥形的,代表生物有竹节石等。
软舌螺纲
软舌螺纲是一个已经灭绝的生物纲,生存年代在寒武纪到奥陶纪,它们的身体呈两侧对称,具有单锥形的壳,代表生物有软舌螺等。
习性
软体动物的习性因种类而异;腹足类在陆地、淡水和海洋均有分布,双壳类只生活在淡水和海洋中,其他类群基本上生活在海洋中。
软舌螺动物门界: 动物界 Animalia
亚界: 后生动物亚界 Metazoa
超门: 冠轮动物总门 Lophotrochozoa
门: 软体动物门 Mollusca
纲: 软舌螺动物门 Hyolitha
软舌螺动物门(学名:Hyolitha)是动物界的一个门,是一类已经灭绝了的海生有壳的无无脊椎动物,其化石一般保存锥壳,口盖和附肢三个部分,外壳为钙质成分,两侧对称。
对于这一类动物的分类上,目前众说纷纭,一种意见认为它是软体动物门之下的一个纲,另一种意见则认为它与软件动无本质上的联系,应属一个独立的门。
人们依据有无口唇将软舌螺动物门(纲)动物分成两个纲(目):
直管螺纲
有唇软舌螺纲
竹节石域: 真核生物域 Eukaryota
界: 动物界 Animalia
门: 软体动物门
纲: 竹节石
目: 竹节石目(Tentaculitida)
等环节石目(Homoctenida)
珠胚节石目(Dacryoconarida)
竹节石是一类已经绝灭的海生无脊椎动物,其归类尚未明确,目前通常根据竹节石化石的壳壁结构和成分以及壳体形态,将该类动物作为软体动物门中的一纲。
根据竹节石壳壁的厚度、初房形态和有无隔壁将竹节石纲划分为3个目:
竹节石目(Tentaculitida)
等环节石目(Homoctenida)
珠胚节石目(Dacryoconarida)
太阳女神螺纲太阳女神螺纲(Helcionelloida)是一类已灭绝的古老软体动物。太阳女神螺纲是已知最古老的有壳亚门生物;也就是说这类生物有一个矿化外壳。其下部分物种曾被分类为单板纲。太阳女神螺纲由John S Peel 于1991年建立。
喙壳纲域: 真核域 Eukarya
界: 动物界 Animalia
亚界: 真后生动物亚界 Eumetazoa
(未分级) 两侧对称动物 Bilateria
原口动物 Protostomia
超门: 冠轮动物总门 Lophotrochozoa
门: 软体动物门 Mollusca
纲: 喙壳纲 Rostroconchia
目
利培壳目 Ribeirioida
强壮壳目 Ischyrinioida
锥鸟壳目 Conocardioida
喙壳纲(学名:Rostroconchia)是软体动物门中的一个纲,这一纲动物目前末发现有生种,全部为化石种。本纲动物的形态接近双壳类,所以在发现之初,曾以为是同类物种。但现时已知它们的壳体最初阶段是从一个原点形成一个原壳,再发育成为具有双壳形态的成年壳体。因此,这是一类具有“假双壳”形态的单壳软体动物
本纲动物从寒武纪早期出现,到二叠纪晚期绝灭,1972年被正式命名为喙壳纲。
目前全世界发现的喙壳纲动物化石共有40多个属,综合其外部形态、内部构造和个体发育等特征,将其分为3目7科:
利培壳目(Ribeirioida) 利培壳科(Ribeiriidae)
敏蜂壳科(Technophoridae)
强壮壳目(Ischyrinioida) 强壮壳科(Ischyriniidae)
锥鸟壳目(Conocardioida) 始翼壳超科(Eopteriacea) 始翼壳科(Eopteriidae)
锥鸟壳超科(Conocardiacea) 锥鸟壳科(Conocardiidae)
勃氏壳科(Bransoniidae)
马心壳科(Hippocardiidae)
单板纲域: 真核生物域 Eukaryota
界: 动物界 Animalia
亚界: 真后生动物亚界 Eumetazoa
(未分级) 两侧对称动物 Bilateria
原口动物 Protostomia
超门: 冠轮动物总门 Lophotrochozoa
门: 软体动物门 Mollusca
目
罩螺目(Tryblidioidea)
古祐目(Archinacelloidea)
弓壳目(Cyrtonellida)
马哈螺目(Maikhanellidae)
Pelagiellida
Tergomya
单板纲(学名:Monoplacophora),又名新蝶贝纲,是软体动物门的一个多系群纲,有一个帽子形状的外壳,在深海的底层生活。单板纲大多数是化石种类,主要产于早古生代海相地层中。现存7种。1952年在南美洲的深海中首次发现该类动物生种,被命名为均为加拉提亚新蝶贝,后来又陆续在其他海域发现了一些生种,它们的内部器官有假分节现象,这为许多学者关于软体动物与环节动物同源的说法。
虽然许多单板纲的外壳有着帽贝的外形,他们并不是腹足类,而且也没有密切关连。
本纲动物的分类有比较大争议,同一物种,在不同的参考可能会被归入不同的分类。以下为比较常见的分类:
罩螺目 Tryblidioidea Pilsbry, in Zittel-Eastman, 1899
罩螺总科 Tryblidiidae von Zittel, 1899
弓壳科 Cyrtonellidae
Family Laevipilinidae Moskalev et al., 1983
单板科 Monoplacophoridae Moskalev et al., 1983
新蝶贝科 Neopilinidae Knight & Yochelson, 1958
罩螺科 Tryblidiidae Pilsbry, in Zittel-Eastman, 1899
Family VemidaeMoskalev et al., 1983
帐篷螺目 Scenellacea
新月贝科 Neomenioida
窄套目 Stenothecoidacea
地位未定
Family Palaeacmaeidae
根据布歇特和洛克罗伊的腹足类分类(2005年)亦包括古生代地位未定软体动物,因为这些物种未能分辨到底是腹足纲还是单板纲。
多板纲界: 动物界 Animalia
门: 软体动物门 Mollusca
亚门: 双神经亚门 Aculifera
纲: 多板纲 Polyplacophora
多板纲(学名:Polyplacophora)是软体动物门的一个纲。本纲物种生活于海中,当中大部分的物种生活在潮间带和潮下带浅水区的岩石上,但也有一些物种类生活在较深海域。其特征为,内圈一般都有八片紧邻排列之硬壳,外圈则是肌肉组织。多板纲包括各种石鳖,常可在退潮后的潮间带岩石上见到。可以食用。
现时本纲物种的分类基本上依从Sirenko (2006)的分类,大致如下:
多板纲 Polyplacophora de Blainville, 1816
古石鳖亚纲 Paleoloricata Bergenhayn, 1955
Order Chelodida Bergenhayn, 1943
Order Septemchitonida Bergenhayn, 1955
新石鳖亚纲 Neoloricata Shumacher, 1817
鳞侧石鳖目 Lepidopleurida Thiele, 1910:例如鳞侧石鳖(Lepidopleurus)。
石鳖目 Chitonida Thiele, 1910:包括多板纲中绝大多数的种类,例如:花斑锉石鳖。
毛肤石鳖亚目 :体呈长形或椭圆形,大小不一。例如:红条毛肤石鳖
目地位未定(Incertae sedis)
Family Scanochitonidae Bergenhayn, 1955
Family Olingechitonidae Starobogatov et Sirenko, 1977
Family Haeggochitonidae Sirenko et Starobogatov, 1977
Family Ivoechitonidae Sirenko et Starobogatov, 1977
无板纲域: 真核域 Eukarya
界: 动物界 Animalia
亚界: 真后生动物亚界 Eumetazoa
(未分级) 两侧对称动物 Bilateria
原口动物 Protostomia
超门: 冠轮动物总门 Lophotrochozoa
门: 软体动物门 Mollusca
纲: 无板纲 Aplacophora
无板纲(学名:Aplacophora)是软体动物门的一个纲。仅有250种左右。是一类身体呈蠕虫状的软体动物。
过往当生物分类学纯綷以形态学为分类准则之时,无板纲物种曾被以为跟海参同类,这一点可在1987年之前的分类中看见。从1987年起,本分类得到确认为软体动物之一,因而成为一个独立的纲。当时本纲由两个群组成,分别为:
沟腹亚纲(Solenogastres):亦作新月贝目,WoRMS把本分类提至纲级。
尾腔亚纲(Caudofoveata):亦作毛皮贝目,同样地WoRMS亦把本分类提至纲级
这两个群最初以为只是并系群,但近年的分子数据支持两个群是单系群的论说。
从分子数据及化石的分析,无板纲很大可能是从多板纲演化,只是后来这些“板”都退化了。
腹足纲界: 动物界 Animalia
门: 软体动物门 Mollusca
纲: 腹足纲 Gastropoda
Cuvier, 1797
亚纲: 直腹足亚纲、 腹足亚纲
腹足纲(学名:Gastropoda)是软体动物门中重要的组成部分,包括通常所说的蜗牛、海螺和蛞蝓。是软体动物门中最大的纲,约60,000到80,000生存种。包括约409个现生的科,另外还有约202个化石科。是动物中仅次于昆虫纲(Insecta)的第二大纲。
头部发达,腹面有肥厚而广阔的足,所以得名;身体有内脏的部分扭转,因此左右不对称;外面有介壳一枚或无壳。
生活在海水中的被称为海螺或海蛞蝓,生活在淡水中的被称为田螺或螺蛳,以及生活在陆地上的蜗牛和蛞蝓。
绝大多数的腹足纲的物种都具有一个呈螺旋形的“壳”,当遇到危险的时候会将柔软的身体缩进壳中。也有一些壳退化消失或仅在体内残留,通常被称为蛞蝓。
腹足纲在全世界范围内分布,包括海洋(约30,000种)、淡水(约5,000种)或陆地(约30,000种)等,从靠近南极和北极的寒冷地区到热带地区都有分布。
可以适应非常多样的生存环境,从花园、森林、沙漠到山区,从沟渠、河流到湖泊,从河口、泥滩、布满岩石的潮间带、沙底的潮下带到深海,还有些营寄生。
双壳纲界: 动物界 Animalia
门: 软体动物门 Mollusca
纲: 双壳纲 Bivalvia
Linnaeus, 1758
亚纲
古列齿亚纲 (Palaeotaxodonta)
隐齿亚纲 (Cryptodonta)
翼形亚纲 (Pteriomorphia)
古异齿亚纲 (Palaeoheterodonta)
异齿亚纲 (Heterodonta)
异韧带亚纲 (Anomalodesmata)
双壳纲(学名:Bivalvia),旧作瓣鳃类(Lamellibranchia)或斧足类(Pelecypoda),是软体动物门的一个纲,生于海洋及淡水地区因有两片贝壳而得名。有7500种左右,体长达135厘米。本纲动物的头部已经退化,足部呈斧状,体躯两侧各有1对瓣状的鳃。壳侧生,开的过程是被动的,其关闭则需要相关肌肉的收缩完成。纤毛抖动在腮部扬起漩涡,使得水及其中的颗粒进入腮部。口通过一条粘膜道以及触须吸取营养颗粒
双壳类软体动物的许多种是重要的养殖和捕捞对象,如珍珠贝等。
在2010年5月,一个有关双壳纲的新分类在期刊《Malacologia》发表。在这篇论文中,作者使用了多种系统发生学的资讯,例如:分子分析学,解剖分析学,外壳形态学及外壳微构造等方法,去把本纲的物种重新分类。根据这个新的分类,双壳纲的物种可以分成324个有效的科,当中有214个科只有化石样本,110个物种延续至现在。这个分类已为WoRMS接纳。
掘足纲域: 真核域 Eukarya
界: 动物界 Animalia
亚界: 真后生动物亚界 Eumetazoa
(未分级) 两侧对称动物 Bilateria
原口动物 Protostomia
超门: 冠轮动物总门 Lophotrochozoa
门: 软体动物门 Mollusca
纲: 掘足纲 Scaphopoda
Bronn, 1862
目
角贝目
管角贝目
掘足纲是软体动物门的一个纲。全部是海产泥沙中穴居的一类小型软体动物,仅有350种左右。海生,直至7000米可见其踪迹。
贝壳长,似象牙,管状,成细弯刀型,两端开口。体长达14厘米。触手从下端开口伸出,不断开挖沉积物将身体大部分埋藏于沉积物中。通过触手将蓝藻,碎石等物质送到口部。上端开口保持其与外界接触。
有重力感觉器,无眼,但触手部分有感觉细胞。
目前仍不能肯定,它们是否具有心脏及其包膜。
头足纲界: 动物界 Animalia
门: 软体动物门 Mollusca
纲: 头足纲 Cephalopoda
Cuvier, 1797
亚纲
鹦鹉螺亚纲(Nautiloidea)
菊石亚纲(Ammonoidea)
蛸亚纲(Coleoidea)
新蛸亚纲(Neocoleoidea)
头足纲(学名:Cephalopoda)是软体动物门的一个纲。化石种在一万种以上,现仅存786种,主要是各类乌贼和章鱼。头足纲动物为全部海生,肉食性,身体两侧对称,分头、足、躯干三部分。头部发达,两侧有一对发达的眼。足着生于头部,特化为腕和漏斗,故称头足类。漏斗位于头部腹面,在头和躯干之间。原始种类具有外壳,现存种类则多是内壳或无壳。鳃为羽状,一对或二对,心耳和肾的数目和鳃一致。口腔具有颚片和齿舌。神经系统集中,感官发达。循环系统为闭管式。直接发育(无需变态)。
头足纲可分为两个到四个亚纲,其中现存两个亚纲。一个是蛸亚纲(Coleoidea)又称为二鳃亚纲,外壳已经消失或是内化。此亚纲包括章鱼、乌贼、鱿鱼等。另一个是鹦鹉螺亚纲(Nautiloidea)又称为四鳃亚纲,外壳依然存在,此纲包括鹦鹉螺等。另有已灭绝的菊石亚纲(Ammonoidea,也可以分类为四鳃亚纲)和箭石亚纲(Belemnoidea)。
头足纲分布在所有海域的所有深度,目前没有发现适应淡水的种类,但有些能够适应不同盐度的水。
神经系统与行为
头足类的神经系统发达,由中枢神经系统,周围神经系统和交感神经系统组成,结构复杂。
头足纲被认为是最聪明的无脊椎动物,因为它们有高度发展的知觉和较大的脑。它们的脑比腹足纲和双壳纲都来的大。除了鹦鹉螺之外,头足纲的表皮拥有一种特殊的色素细胞(chromatophore),使他们能够经由变色来进行沟通和伪装。有研究显示一只章鱼的智力有如五岁的孩童,能够进行开罐、玩积木等较复杂的动作。
此外头足纲的神经系统是无脊椎动物之中最为复杂的,在外套膜(mantle)中庞大的神经纤维成为神经生理学常用的实验材料。
头足纲的视觉敏锐,实验证明普通的章鱼能够辨识亮度、形状、大小还有物体的垂直和水平方向。头足纲的眼睛更能够感应光线的偏振平面。令人惊讶的是,这些能够变色的头足纲动物大都是色盲。当他们进行伪装的时候,能够依照他们所看见的背景,利用色素细胞改变皮肤的亮度和花纹。而改变颜色的时候使用的是彩虹色素细胞(iridophore)和白色素细胞(leucophores),这些细胞能够反映环境的光线。目前为止,只有一个种类的彩色视觉(color vision)得到证明,称为萤火鱿(Sparkling Enope Squid)。
循环系统
头足纲是软体动物当中唯一拥有封闭式循环系统的分类:它们拥有两个经由鳃中的微血管来输送血液的鳃心;一个经由身体的其他部分输送充氧血的单一系统心脏(single systemic heart)。
和其他的软体动物一样,头足纲利用一种含铜离子的血青蛋白(hemocyanin)来运送氧气,而不是像鸟类或一般哺乳动物使用血红素。它们的血液缺氧时呈无色透明;接触空气之后时呈蓝色。
运动方式
头足纲的一般行动方式是利用喷射动力,充满氧气的水被吸入外套膜中的鳃之后,肌肉收缩使空间减少,导致水从由足演变而成的漏斗管(hyponome)喷出,通常是背对着水喷出,并且能够用漏斗控制方向。这是一种相对用尾巴推进更为耗能的移动方式,相对效率随着体型增大而降低,这也使一些种类尽可能使用鳍和臂来推进。
有一些种类的章鱼能够在海底行走,墨鱼和乌贼可以摆动外套膜上的翼状肌肉来移动。
繁殖与生命周期
除了少数例外,蛸亚纲的寿命很短且成长快速,大多数吃下的养分都被它们用来长大。大多数种类的雄性阴茎是一个用来将精囊输送到交接腕(hectocotylus)的生殖管(gonoduct)肌肉末端。而交接腕是用来将精囊输送给雌性。有一些种类没有交接腕,它们直接将较长的阴茎伸出外套膜来与雌性直接交配。此外他们行"单次繁殖",也就是一生只生产一次。下完一整组的蛋之后便死亡。
而鹦鹉螺亚纲则行“多次繁殖”,它们寿命较长,且一次只下少量的蛋。
演化
头足纲在寒武纪晚期出现,在古生代和中生代期间支配并分化出水生型态。已灭绝的托莫特壳类是头足纲的原始型态,它有类似章鱼的触腕,但却用类似蜗牛的腹足来行走于海床。早期的头足纲位在食物链的顶端。
不论是古代的箭石类(Belemnoidea)或是现代的新头足类(Neocoleoidea),或是菊石(ammonoids),都是由拥有外壳的鹦鹉螺类在4.5亿年前到3亿年前的古生代分化出来。古代的头足纲有外壳保护,这些外壳原本是圆锥状,但是后来变成了鹦鹉螺那样的螺旋形状。到了现代,许多的种类依然有内壳,而大多数拥有外壳的种类在白垩纪就消失了。
分类
头足纲
鹦鹉螺亚纲(Nautiloidea):又称四鳃亚纲
短棒角石目(Plectronocerida):头足纲在寒武纪的祖先
爱丽斯木角石目(Ellesmerocerida):包括所有之后头足纲的祖先
内角石目(Endocerida)
珠角石目(Actinocerida)
叠盘角石目(Discosorida)
假直角石目(Pseudorthocerida)
塔飞角石目(Tarphycerida)
箭钩角石目(Oncocerida)
鹦鹉螺目(Nautilida):鹦鹉螺和化石亲戚,如珍珠鹦鹉螺
袋角石目(Ascocerida)
直角石目(Orthocerida)
杆石目(Bactritida):包括所有蛸亚纲和菊石亚纲祖先
菊石亚纲(Ammonoidea):已灭绝,也有分类方法把菊石归入四鳃亚纲
棱菊石目(Goniatitida)
齿菊石目(Ceratitida)
菊石目(Ammonitida)
蛸亚纲(Coleoidea)
箭石目(Belemnoidea):已灭绝
沟箭石目(Aulacocerida)
闭箭石目(Phragmoteuthida)
Hematitida
真箭石目(Belemnitida)
新蛸亚纲(Neocoleoidea):也称为二鳃亚纲
十腕总目(Decapodiformes)
旋乌贼目(Spirulida):Ram's Horn Squid
墨鱼目(Sepiida):墨鱼
耳乌贼目(Sepiolida):大西洋耳乌贼
管鱿目(Teuthida):鱿鱼、章鱿、大王乌贼
八腕总目(Octopodiformes)
吸血鬼乌贼目(Vampyromorphida):活化石,又称幽灵蛸
章鱼目(Octopoda):章鱼(Octopus)、船蛸(Argonauta)、水母蛸(Amphitretus)等
鹦鹉螺界: 动物界 Animalia
门: 软体动物门 Mollusca
纲: 头足纲 Cephalopoda
亚纲: 鹦鹉螺亚纲 Nautiloidea
目: 鹦鹉螺目 Nautilida
亚目: 鹦鹉螺亚目 Nautilina
Agassiz, 1847
科: 鹦鹉螺科 Nautilidae
Blainville, 1825
属/种
异鹦鹉螺属(Allonautilus)
穿孔异鹦鹉螺(Allonautilus perforatus)
异鹦鹉螺(Allonautilus scrobiculatus)
鹦鹉螺属(Nautilus)
帕劳鹦鹉螺(Nautilus belauensis)
Nautilus clarkanus
可可尔鹦鹉螺(Nautilus cookanum)
大脐鹦鹉螺(Nautilus macromphalus)
珍珠鹦鹉螺(Nautilus pompilius)
Nautilus pompilius pompilius
Nautilus pompilius suluensis
正鹦鹉螺(Nautilus praepompilius)
白斑鹦鹉螺(窄鹦鹉螺)(Nautilus stenomphalus )
鹦鹉螺,海洋软体动物,仅存于印度洋和太平洋海区,北至日本南方,南至大堡礁,西至安达曼海,东至斐济等地区均有发现。位于鹦鹉螺主要产地的法属新喀里多尼亚,还以鹦鹉螺做为国徽的主要图案。鹦鹉螺是四大名螺之一。
现存种群数量少。因生活环境须有数个大气压的水压,故难人工饲养。
鹦鹉螺已经在地球上经历了数亿年的演变,但外形、习性等变化很小,被称作海洋中的“活化石”,在研究生物进化和古生物学等方面有很高的价值。
在帕劳共和国内的深海中尚有鹦鹉螺的栖息地。
鹦鹉螺外形色彩绚丽,体型华美,观赏性极强。它的壳薄而轻,呈螺旋形盘卷,壳的表面呈白色或者乳白色,生长纹从壳的脐部辐射而出,平滑细密,多为红褐色。整个螺旋形外壳光滑如圆盘状,形似鹦鹉嘴,故此得名“鹦鹉螺”。
鹦鹉螺外壳切面呈现优美的等角螺线,而等角螺线本身又与黄金分割有着密不可分的关系,这使鹦鹉螺在亮丽的外表之外又增加了许多大自然的神秘色彩。
鹦鹉螺外形与普通头足纲动物类似,有显著的头部和触须。但鹦鹉螺的触须明显偏多,最多可达90根。鹦鹉螺的触须没有吸管,相互之间并无功能上的区别,而且可以收回。鹦鹉螺的齿舌很大,并有9颗牙齿,两对鳃。
在现在存活的头足纲动物中,仅有鹦鹉螺具有贝壳,其他头足纲动物均无贝壳;船蛸这类的章鱼虽然也有壳,但那只有母船蛸在产卵时会产生白色系的外壳以保护卵,而且外壳与肉体可分离,产卵后便不再有外壳,然而鹦鹉螺的肉体与贝壳是不会分离的。
鹦鹉螺的贝壳最大可为26.8公分,但成年鹦鹉螺一般都不超过20公分。深脐鹦鹉螺(Nautilus macromphalus)是鹦鹉螺家族中体形最小的属种,一般只有16公分。
鹦鹉螺的壳被多个横断的隔板分隔成三十余个独立的壳室,除动物体所在的最后一个大壳室外,都充满气体(多为氮气),当动物体不断成长,房室也周期性向外侧推进,在外套膜后方则分泌碳酸钙与有机物质,建构起一个崭新的隔板。各壳室之间有一个贯穿的细管,用以输送气体进到各壳室之中,通过气体的调节,操纵身体浮沉与移行。鹦鹉螺这种特殊的身体结构也为人类建造潜艇提供了灵感,许多国家的潜艇也以“鹦鹉螺”命名。
鹦鹉螺为暖水性动物,生活适宜水温为19-20℃,一般生活在50到300米深的海洋中,通常夜间活跃,多以底栖的小蟹、小虾等甲壳类动物幼体为食,日间则躲在珊瑚礁浅海的岩缝中,以触手握在岩石上歇息,当动物死亡后,身躯软体脱壳而沉没,其空壳则随洋流漂移,研究其漂移路线对洋流的分析有一定意义。
鹦鹉螺雌雄异体,卵生,雌鹦鹉螺每年产卵一次,一般将卵产于浅水岩石上,孵化期12个月,新出壳的小鹦鹉螺一般体长约3公分。
鹦鹉螺寿命一般为二十年,是头足纲动物中寿命较长的动物。
名称和形象的使用
鹦鹉螺号:科幻小说家儒勒·凡尔纳的代表作之一《海底两万里》中虚构的潜艇;
鹦鹉螺号:世界上第一艘实际运作服役的核动力潜艇;
鹦鹉螺:在虚空派画作中代表完美
菊石亚纲界: 动物界 Animalia
门: 软体动物门 Mollusca
纲: 头足纲 Cephalopoda
亚纲: 菊石亚纲 Ammonoidea
Zittel, 1884
目
菊石目 Ammonitida
棱菊石目 Goniatitida
齿菊石目 Ceratitida
菊石亚纲(学名:Ammonoidea)是一群已经灭绝的海洋生物总称,非常适合作为标准化石,地质学家可以使用它们来确定含有菊石化石的地层的年代。菊石亚纲与现存的头足纲关系最接近的可能是蛸亚纲,而不是鹦鹉螺亚纲的鹦鹉螺目。
菊石亚纲约在志留纪晚期至泥盆纪初期第一次出现在地球上,最后与恐龙一起于白垩纪晚期灭绝。
生物学家根据化石的壳室的结构与外壳的花纹,目前将菊石亚纲分成3个目:菊石目、棱菊石目与齿菊石目。其中菊石目分成五个已知的亚目。
菊石的壳沿平面卷曲,呈盘状,两面对称,壳表面光滑或具细的生长线纹,有些具特殊的纹饰,如纵棱、横肋、瘤和刺等。
菊石一般漂浮在海水上层,下面经常是极其缺氧区域,没有生物。菊石死后,沉到海底,逐渐埋没。细菌分解遗体时候,把附近水中性质变化,降低矿物质溶解度,尤其是磷酸盐和碳酸盐。菊石化石上面有一圈一圈的矿物质,因此保存很多高质标本。
棱菊石目界: 动物界 Animalia
门: 软体动物门 Mollusca
纲: 头足纲 Cephalopoda
亚纲: 菊石亚纲 Ammonoidea
目: 棱菊石目 Goniatitida
Hyatt, 1884
亚目
棱菊石亚目
圆叶菊石亚目 Tornoceratina
Anarcestina
Clymeniina
棱菊石目(Goniatitida)是一群已经灭绝的菊石亚纲生物,与乌贼、章鱼关系较接近,与鹦鹉螺关系较远。棱菊石目最早出现在泥盆纪早期(大约4亿年前),兴盛于石炭纪与二叠纪,最后于古生代末期,约2亿4500万年前灭绝。 所有的棱菊石类都拥有外壳,硬壳内部被隔板分隔成独立的壳室,壳室里可能都充满着气体。大部分的棱菊石类直径都小于15公分,甚至常常小于5公分。棱菊石类的外壳通常都具有螺旋状的花纹,与中生代的菊石类不同。中生代的菊石类外壳大部分只有部分螺旋花纹,有些种类甚至没有螺旋花纹。
菊石目界: 动物界 Animalia
门: 软体动物门 Mollusca
纲: 头足纲 Cephalopoda
亚纲: 菊石亚纲 Ammonoidea
目: 菊石目 Ammonitida
Hyatt, 1889
亚目
菊石亚目 Ammonitina
勾菊石亚目 Ancyloceratina
叶菊石亚目 Phylloceratina
弛菊石亚目 Lytoceratina
菊石目(学名Ammonitida)是一类已灭绝的头足类。它们是非常好的指准化石,往往能连结起所在的地层与地质年代。菊石目的近亲并非现今外观相似的鹦鹉螺,反而是八爪鱼、乌贼及鱿鱼等鞘亚纲。
菊石目的化石壳一般呈平旋状,但也有呈垂直螺旋状及非螺旋状。老普林尼称菊石目为“阿蒙的角”,因其壳的形状像埃及神话阿蒙所配带的羊角,其学名亦是以此来取的。其下的属亦多以希腊文“角”的字根来结尾。
菊石目下已命名了四个亚目,分别为菊石亚目(下侏罗纪至上白垩纪)、勾菊石亚目(上侏罗纪至上白垩纪)、叶菊石亚目(下三叠纪至上白垩纪)及弛菊石亚目(下侏罗纪至上白垩纪)。不过《古无脊椎动物学论丛》(The Treatise on Invertebrate Paleontology)却只包括了菊石亚目、弛菊石亚目及叶菊石亚目,并将之分类在菊石亚纲之下;而勾菊石亚目则被看为弛菊石亚目下的一个超科。另外有学者将这三个亚目分类在菊石目中。
根据论丛所述,菊石亚目是于侏罗纪初期由叶菊石亚目及弛菊石亚目衍生而来。
菊石目已灭绝,保存下来的软体部分亦甚为稀有。大部分研究都是基于菊石目的壳及壳的模型。
大部分菊石目都可能生活在远古海洋的开放水域,而非海底。这是由于它们的化石往往都是在没有底栖动物的地层发现。很多菊石目相信都是游泳能手,如锐棱菊石,因它们的壳扁平、呈铁饼状及流线形,但也有一些可能是慢游或底栖的。菊石目及其亲属可能会猎食鱼类、甲壳类及其他细小生物;它们的天敌包括水生爬行类,如沧龙科,在化石化的菊石目都有一些牙齿痕迹。它们可能会喷墨汁来逃避掠食,因为一些化石标本保留了这些墨汁。
菊石目的软体部分占有了壳末的最大节。较早而细小的节都有间隔,可以透过填充气体来维持浮力。故此较细小的部分会浮在较大的之上。
菊石目壳有间隔的部分称为闭锥。闭锥包含了逐渐变大的住室,住室之间有一道很薄的间隔隔开。菊石目动物只住在最后及最大的住室中。当它渐长时,会在开口处增加较新及较大的住室。连室细管由菊石目的身体,穿过间隔至所有空置的住室。利用主导的高渗透运输作用,菊石目可以将这些住室的水份抽走,从而控制壳的浮力在水柱中上升下沉。
菊石目与鹦鹉螺类主要不同之处是在于其连室细管在沿腹面穿过间隔的,而鹦鹉螺类的则是位于接近中央位置。
现今鹦鹉螺壳的大小会与性别有关:雄性的壳较细小,而雌性的壳较大。这种两性异形可能也在菊石目中出现,亦正好解释了它们大小的变异。雌性需要较大的壳可能是为产卵之用。侏罗纪早期欧洲的对菊石就是其中一个例子。菊石目壳的两性异形只是近年的发现。以往这种大小的差异被认为是互为近亲的不同物种。
大部分菊石目壳都是呈扁平旋状,但也有一些是接近笔直的(如杆菊石)、像腹足纲壳的螺旋(如塔菊石及伯斯特菊石)、部分展开及卷曲而成熟时是笔直的(如澳洲菊石)。部分及完全展开的形态于白垩纪初期开始变得多样化,并被称为“异形菊石”。
异形菊石中外观最极端及异常的算是日本菊石,它们就像是一团不规则的螺旋,没有明显对称的卷曲。不过当近看时,就可以见到日本菊石壳上有U形的三维网络。
菊石目壳的表面分野很大。一些表面除了生长线外就很光滑,就像是现今的鹦鹉螺;一些则可以有螺旋脊及肋骨,甚至有棘伸出。这类的菊石目属白垩纪较后期出现。
一些菊石目是连同一块角状板或一对方解石板一同被发现。以往认为这些板是用来开合其壳,就像是鳃盖一样。不过近年却认为这其实是其颚部。这些板统称为口盖,单一块的称为单口盖,成对的则是双瓣颚。不论是单口盖或双瓣颚都很少能保存下来,而且都是与壳分离。单口盖通常都是属于泥盆纪至白垩纪的菊石目,而双瓣颚则是属于中生代的菊石目。双瓣颚是对称的,大小及外观一样。
口盖有多种形状,内外表面的纹刻也各有不同。但是,由于它们很稀有,故至今仍不清楚哪一类口盖是属于哪一个物种,一些口盖甚至被立为独立的属。
虽然菊石目的化石数量很多,但其软体却十分稀少。除了一些墨汁囊及可能的消化器官外,并没有发现任何软件部分。估计它们有很多触手来捕捉猎物。
侏罗纪早及中期的菊石目很少会超越直径23厘米的大小。到了侏罗纪晚期及白垩纪早期,就开始出现了一些较大的形体,如英格兰南部的泰坦菊石直径就达53厘米,而白垩纪德国的Parapuzosia seppenradensis就更有达2米的直径。北美洲已知最大的菊石目就是Parapuzosia bradyi,直径长1.37米。另外,在英属哥伦比亚发现了直径达2.3米的未确认种,一旦获得确认,就是已知最大的菊石目。
由于菊石目可能是游泳或浮游的,它们往往都是栖息在海床以上的水域。当它们死亡后,就会下沉到海床及埋在沉积下,尸体会被细菌所腐化来维持平衡。很自然地,其化石周边就会出现矿物质的同心沉淀,称为结核,也是可以好好保存菊石目化石的原因。
在黏土中发现的菊石目表面都会保存了珠母层的涂层,如英格兰的盔菊石。
美国及加拿大的白垩纪皮埃尔泥板岩出产大量的菊石目化石,包括杆菊石、饼菊石、船菊石、装甲船菊石及Jeletzkytes,与及大量展开的形态。这些化石大部分都能保存原有的壳。
其他化石,如在马达加斯加及加拿大艾伯塔省发现的,都会有虹色。这些虹色菊石目在打磨后都有宝石的质素。相信这种虹色在活生生的菊石目并不存在,而是在其壳上多了额外的一层。
于中生代以后,菊石目化石就开始减少,直到新世代就完全消失。最后的生存线是于6500万年前的白垩纪-第三纪灭绝事件。于白垩纪晚期的生物集群灭绝后就没有菊石目生存,但却因个体发生学的分野而存留了一些鹦鹉螺类。若菊石目的灭绝是因火球撞击,周边的浮游生物会严重消失,造成菊石目繁殖的灾难。
于中世纪的欧洲,菊石目化石被认为是石化了的蛇,故又被称为蛇石。相传这是由怀比特的希尔达(Hilda of Whitby)或圣帕特里克等圣人事迹的证据。商人会在菊石目的开口刻上蛇的面。菊石目在尼泊尔卡格里达金河(Gandaki river)被称为“萨里格拉姆”,印度教相信它们是毗湿奴的显现。
勾菊石亚目界: 动物界 Animalia
门: 软体动物门 Mollusca
纲: 头足纲 Cephalopoda
亚纲: 菊石亚纲 Ammonoidea
目: 菊石目 Ammonitida
亚目: 勾菊石亚目 Ancyloceratina
Wiedmann, 1966
超科
Ancylocerataceae
Douvilleicerataceae
塔菊石超科 Turrilitaceae
德沙耶斯菊石超科 Deshayesitaceae
Scaphitaceae
勾菊石亚目(学名Ancyloceratina)是一类多样化的菊石,菊石目下弛菊石亚目的近亲。它们于侏罗纪晚期开始演化,至白垩纪期间非常普遍及多样化。它们遍布各大洲,可以作为指准化石。大部分勾菊石亚目的壳都不是螺旋状,故属于异形菊石。
较原始的勾菊石亚目的形状接近完整的螺旋,只是在最末及外端开放,像螺旋下的一个钩。以这种形状而言,螺旋的部分就是浮起的部分,而钩就是住室。例子包括钩菊石、Protanisoceras及Tropaeum。较后期的形态与之有甚大的差别。例如Ptychoceras的壳就有三或四个挤在一起及互相连接的筒。钩角石的形状就像是一个回形针。
到了白垩纪晚期,它们的形状更为多变,甚至有完全笔直的杆菊石及Sciponoceras、扭转螺旋状的塔菊石、并奇形怪状的日本菊石。一些形态是综合了多个形状,例如Nostoceras,开始是像塔菊石的螺旋形,底下是像钩菊石较水平的钩。
并非所有勾菊石亚目的形状都如此特殊的。很多早期的形态甚至很难于普遍的菊石目分辨出来。一些展开的分支后来也取回螺旋的形状,最著名的包括船菊石、装甲船菊石及其亲属。
勾菊石亚目的生态不明,但有一点可以肯定的是展开形态的并不适合游泳。一些拥有开放式壳的很易制造旋涡,而其壳也是浮在软件之上,对游泳造成极大的阻碍。所以,它们很有可能只是浮游的,以长触手来捕捉细小猎物;又或是底栖的。
勾菊石亚目的大小差异很大:最细小的Ptychoceras只有约3厘米长;而杆菊石及Diplomoceras就可以长达1-2米。一些物种如钩角石就广泛分布在欧亚大陆、南美洲及南极洲;而另一些就可能只限于西部内陆海道。
箭石目界: 动物界 Animalia
门: 软体动物门 Mollusca
纲: 头足纲 Cephalopoda
亚纲: 蛸亚纲 Coleoidea
下纲: 箭石下纲 Belemnoidea
目: 箭石目 Belemnitida
Zittel, 1895
箭石目(学名:Belemnitida)是一种已经灭绝的头足纲生物,生活在泥盆纪至白垩纪之间。箭石在许多方面都与现代的鱿鱼相当接近,而且与现代的乌贼关系密切。箭石也拥有墨汁,不过它们有10条大约相同长度的触腕,其中并没有特别长的触腕。
箭石在侏罗纪与白垩纪时数量相当多,所以它们的化石在中生代的海洋岩层中相当丰富,经常伴随着菊石类出现。不过箭石也跟菊石一起在白垩纪末期灭绝了。
现时箭石目只包含下列三个亚目:
Belemnitina
Belemnopseina
Belemnotheutina
以下三个亚目原来属于箭石目,现时已分往其他分类:
Aulacocerida
Phragmoteuthida
Diplobelida
蛸亚纲界: 动物界 Animalia
门: 软体动物门 Mollusca
纲: 头足纲 Cephalopoda
亚纲: 蛸亚纲 Coleoidea
Bather, 1888
下纲及目
箭石下纲 Belemnoidea
Aulacocerida
闭箭石目 Phragmoteuthida
Donovaniconida
箭石目 Belemnitida
Hematitida
新蛸下纲 Neocoleoidea
Boletzkyida
乌贼目 Sepiida
耳乌贼目 Sepiolida
旋乌贼目 Spirulida
枪形目 Teuthida
八腕目 Octopoda
幽灵蛸目 Vampyromorphida
蛸亚纲(学名Coleoidea)是头足纲下的一类软体动物。它们不像鹦鹉螺亚纲般有硬壳,最多只有用来控制浮沉的内骨。一些物种甚至完全没有骨头,一些则以软骨来支撑。
蛸亚纲的分类是以触手的数量及结构来定界的。已灭绝及最原始的箭石下纲相信是有10只触手。较现代的物种其中一对触手可能已经进化或退化了。十腕总目的第四对手臂进化成为有吸盘的长触手。八腕总目的第二对手臂有不同的改变,如幽灵蛸目的感应丝状物或八腕目的已完全退化。
演化历史
最古老的蛸亚纲是源自约于3亿3000万年前石炭纪的密西西比纪。也有发现一些属于泥盆纪的化石,但有学者不认同它们是蛸亚纲。
到了石炭纪,蛸亚纲已经有多样的形态。虽然这些类别传统上都分类为箭石下纲,但它们的分别明显显示它们并非近亲。
分类
头足纲
鹦鹉螺亚纲
菊石亚纲
蛸亚纲
箭石下纲
始乌贼属
Hematitida
闭箭石目
Donovaniconida
Aulacocerida
箭石目
新蛸下纲
十腕总目
Boletzkyida
旋乌贼目
乌贼目
耳乌贼目
枪形目
八腕总目
幽灵蛸目
八腕目
口噬目
十腕总目界: 动物界 Animalia
门: 软体动物门 Mollusca
纲: 头足纲 Cephalopoda
亚纲: 鞘亚纲 Coleoidea
超目: 十腕总目 Decapodiformes
目
Boletzkyida
旋乌贼目(Spirulida)
墨鱼目(Sepiida)
耳乌贼目(Sepiolida)
管鱿目(Teuthida)
异名
Decembrachiata
Winckworth, 1932
十腕总目(学名:Decapodiformes)是头足纲蛸亚纲新蛸下纲之下的一类软体动物,新蛸下纲的另一大分类是八腕总目。
与蛸亚纲的其他分类一样,十腕总目的物种同样有10只触手,但其第四对触手演变成为了有吸盘的长触须。
分类
十腕总目 (Decapodiformes)
Boletzkyida
旋乌贼目(Spirulida):Ram's Horn Squid
墨鱼目(Sepiida):墨鱼
耳乌贼目(Sepiolida):大西洋耳乌贼
管鱿目(Teuthida):鱿鱼、章鱿、大王乌贼
八腕总目界: 动物界 Animalia
门: 软体动物门 Mollusca
纲: 头足纲 Cephalopoda
亚纲: 鞘亚纲 Coleoidea
超目: 八腕总目 Octopodiformes
Berthold & Engeser, 1987
目
八腕目 Octopoda
口噬目 Trachyteuthis
幽灵蛸目 Vampyromorphida
异名
Octobrachia Fiorini, 1981
Vampyropoda Boletzky, 1992
八腕总目(学名:Octopodiformes)是头足纲蛸亚纲新蛸下纲之下的一类软体动物,新蛸下纲的另一大分类是十腕总目。
与蛸亚纲的其他分类一样,八腕总目的物种同样有10只触手,但不同物种的第二对手臂有不同的改变,如幽灵蛸目的演变成为感应丝状物,而八腕目的已完全退化。